No.70 - 自己と非自己の科学(2) [本]
前回の、No.69「自己と非自己の科学(1)」に引き続き、故・多田富雄氏の2つの著作が描く免疫システムの紹介です。ここからは、私自身が多田氏の本を読んで強く印象に残った点、ないしは考えた点です。
免疫システムの特徴
多田氏の2冊の著作が描く免疫システムを概観すると、それは幾つかの際だった特徴をもっていることに気づきます。免疫システムの特徴をキーワードで表すと以下のようになると思います。
免疫は、刻々変わる「自己」と「非自己」に対応してシステムのありようを変え、再組織化していきます。この「自己組織化」は、変化する環境に対応して「困難な」目標を達成するべく運命づけられたシステムの必然なのでしょう。
前回の No.69「自己と非自己の科学(1)」の中の「免疫のプロセス」で、多田氏の本からそのまま引用してB細胞とT細胞が絡む免疫の過程を紹介しました。このプロセスは、かなり複雑でまわりくどく、また冗長だと感じられます。もっとシンプルにできないのか。
免疫学の歴史上有名な「クローン選択説」があります。オーストラリアのウイルス学者・バーネットが1957年代に出した免疫のしくみを説明する学説で、現代免疫学の根幹をなす重要な学説です。それをB細胞を念頭において模試的に描いたのが次の図です。
図で左端のA・B・C・D・EはB細胞です。B細胞はそれぞれ抗原と特異的に反応する抗体を持っていて、それは細胞ごとに違う。ある抗原が図の「B細胞・C」の抗体と反応すると「B細胞・C」は分裂をはじめ、同じ遺伝的性質の子孫(=クローン)を増やしていく。それは「B細胞・C」のもつ抗体の大量生産につながり、抗原は駆除される。しかし一部のB細胞は組織内に長く残り、それが記憶細胞となって二次免疫反応を起こす・・・・・・。
バーネットが「クローン選択説」を出したときにはT細胞もB細胞も分かっていませんでした。しかし免疫の仕組みの基本的なところを見事に言い当てている。もうこの図で十分だという気がします。
しかし実際の免疫の仕組みはこのようにシンプルではありません。もっと複雑で冗長です。それは「免疫のプロセス」で引用したように、
というプロセスをたどります(キラーT細胞は省略)。「B細胞の抗体が抗原と反応しただけでB細胞が分裂を開始し抗体が大量生産される、ということはない」のです。なぜこんなにまわりくどくて冗長なのか。
おそらく上図のようなシンプルな仕組みは「あぶない」のだと思います。もし、自己のタンパク質と反応する抗体を持つB細胞があるとすると、その抗体が大量生産され、自己の破壊につながる。免疫の仕組みの冗長さは、それなりの理由があると考えられます。
抗原を特異的に認識したり無力化するのは、T細胞抗原受容体(TCR)と免疫グロブリン(Ig)でした。そして、無限のバリエーションがある抗原を「特異的」に認識する仕組みは、まず遺伝子のランダムな組み替えで予期しえない TCR や Ig が作られ(利根川博士の発見)、そこから自己と反応するものが排除されるというプロセスでした。
免疫システムの根幹のところには、この「ランダム性」があります。非常に無駄が多い仕組みに見えますが、不測の事態(抗原の進入)の備えるにはこれしかないのでしょう。
B細胞が作り出す抗体(免疫グロブリン。Ig)は、抗原を直接に認識します。しかし抗体の大量生産のトリガーを引くのはヘルパーT細胞であり、ヘルパーT細胞が認識するのは、MHCに提示された抗原の断片(9個のアミノ酸からなるペプチド)でした。
ここで誰しも思うのは「わずか9個のアミノ酸で抗原が確実に認識できるのだろうか」という疑問です。タンパク質はふつう数百のアミノ酸から成り、数千のアミノ酸で構成されるタンパク質も稀ではありません。9個で大丈夫なのか。
実際、多田氏の本には次の意味の記述があります(免疫の意味論)。
抗原のタンパク質断片でもMHCの溝に「提示」可能なものは少なく、かつそれが人間のタンパク質と異なっているとは限らない・・・・・・。免疫システムは、ある種の「偶然性」の上に成り立っていることになります。多田氏が書いているように、この免疫システムの「欠陥」をついてくる「非自己」もある。
しかし多くの場合はこれで免疫が成り立っています。つまり、抗原もMHCもT細胞も「数が圧倒的に多い」ということなのでしょう。個々の確率は少なくても、母数が多いと安定的に作動するということがあります。
前回、No.69「自己と非自己の科学(1)」の「免疫のプロセス」に出てきたインターロイキン6(IL-6)というサイトカインがあります。これはヘルパーT細胞が作りだし、B細胞がプラズマ細胞に分化するように誘導する重要なサイトカインでした。しかしIL-6には別の作用もあるのです。多田氏の本によると、たとえば
◆キラーT細胞の分化を促す
◆マクロファージの働きを高める
などです。このあたりは免疫細胞に関係した作用であって、抗原を排除するのに役立つのだなと納得がいきます。ところが、IL-6は免疫とは直接関係のない作用もあるのです。つまり
◆神経細胞の成長を促す
◆骨を破壊する破骨細胞を作り出す
◆造血幹細胞や血小板を増やす
などです。同じ分子(タンパク質)が、こういった多種の作用を兼ねていいのかと思うのですが、それだけではありません。IL-6を作るのはヘルパーT細胞だけでなく、
◆マクロファージ
◆B細胞
◆皮膚の上皮細胞
◆血管の内皮細胞
でも作られる、とあるのですね。
B細胞をプラズマ細胞に分化させるという、免疫システムにとっては決定的に重要なサイトカインが、多数の別の「意味」に使われ、しかも各種の細胞がそのサイトカインを出す・・・・・・。IL-6というサイトカインはきわめて「多義性」があることになります。そしてこの多義性は IL-6 だけではないのです。
こんなことで免疫が成立するのかと心配になりますが、一応成立している。こういった「多義性」を含みつつ複雑な目的を実現するところに、自己組織化がポイントである免疫システムの特徴があるように感じます。
免疫システムは「危うさ」をもったシステムです。たとえば「胎児を母親の免疫システムから守る仕組み」です。前回の No.69「自己と非自己の科学(1)」で、多田氏の本に従って、
という意味のことを書きました。一見これで良いようなのですが、これは非常に「あぶない」仕掛けです。つまり人間には「MHCの発現を抑制する」遺伝子があり、胎盤だけでその遺伝子が働いているというようなことだと想像します。そうだとしたら、もし、がん細胞がその遺伝子を「利用」したらどうなるのか。免疫システムはがん細胞を攻撃できなくなります。事実、多田氏の本には「がん細胞は細胞のMHCを少なくしT細胞の監視からのがれる」という意味のことをが書かれているのです。
そもそもキラーT細胞は自己の細胞を殺す役割の免疫細胞です。もちろん殺すのは「細胞表面のクラス1・MHCに非自己(抗原)の断片を提示している自己の細胞」で、内部になんらかの異変が起きていると想定できる細胞です。ウイルスは自己に潜む非自己、ないしは自己に偽装した非自己なので、ウイルスの駆除までを目的とする免疫システムでは自己の細胞を殺す必要があるのは理解できます。
しかし免疫の仕組みには「偶然性」や「ランダム性」が内在しています。もし間違って本当の自己を提示している細胞を殺してしまったらどうなるのか。そういった反応性をもつキラーT細胞が「自己に反応する免疫細胞を抑制する仕組み(免疫的寛容)」を乗り越えて大増殖したらどうなるのか。免疫システムが自己を攻撃する事態が想像されます。
さらに想像すると、人間のDNAにある遺伝子は全部が活用されていわけでも何でもなく「眠っている」遺伝子がたくさんあることは生命科学の教えることです。その眠っている遺伝子が何かの拍子に発現し、通常では作られないタンパク質が細胞内で生産されるとどうなるのか。人間の免疫系はその細胞(自己)を攻撃することになります。
いろいろと心配の種が尽きない「危うさ」を内包しているのが免疫システムです。そして自己免疫病と総称される病気はまさにこの「危うさ」が現実のものとなった病気だと感じられます。
現代における難病のほとんどは「自己免疫病」です。たとえばバセドー病は甲状腺に反応する抗体ができる免疫反応で起こります。若年性糖尿病はインシュリンを生産する膵臓のβ細胞を自己の免疫系が破壊してしまうことで起こります。人間のほとんどの臓器・部位に対して、そこを攻撃してしまう自己免疫病があると言います。また体内の関節が炎症を起こし萎縮していくリウマチ性関節炎も自己免疫病です。付け加えますと、いわゆるアレルギーは抗原に対する免疫システムの過剰反応が原因です。それによって人間にとっては不要な、時には命にかかわりかねない炎症が起こる。
自己免疫病で最も破壊的なのはSLE(全身性エリテマトーデス)という難病で、これは自己のDNAに対する抗体ができてしまう病気なのです。DNAはすべての細胞にあるわけですから、免疫システムが「すべての自己」を攻撃する。攻撃されて破壊された細胞からはDNAが飛び出し(=抗原!!)、それがまた抗体の生産を促して、悪循環が際限なく続くことになります。
「自己免疫病」を典型とする免疫システムの「危うさ」は、非自己を排除するという極めて困難な目的の達成のために必然的に組み込まれてしまったものだと思いました。花粉症はその「ささやかな」例なのでしょう。
ここで言う「多様性」は、人間が個体ごとに違うという意味での多様性です。
免疫が発動するプロセスは、まず人それぞれに違うMHCが細胞表面にあり、そのMHCに抗原の断片が提示され、その「MHC・抗原断片の複合体」をT細胞が認識する、という過程でした。そこからロジカルに導かれる結論は、
ということです。特定の抗原を認識しやすい人とそうでない人がいる。インフルエンザが流行して学校が閉鎖になっても、まったく病気にならない子もいる。逆にワクチンを接種してもそれが利かずにインフルエンザにすぐかかってしまう子もいる。スギ花粉を吸い込むとアレルギー症状を起こす人もいれば、起こさない人もいる。抗原(病原体)に対する反応は人によって非常に違うわけです。
世界史をみると15世紀のヨーロッパではペストが大流行しました。人口の3分の1が死亡したと言います。3分の1が死亡するという事態は大パニックを起こしたでしょう。しかしペストが大流行しているさなかでケロッとしている人も多いわけです。当時は「細菌による伝染病」という概念は全くありません。死んだ患者と接触した人も多かったはずです。それでも3分の2は死ななかった。もし人口の9割が死んだとしたらヨーロッパ文明は崩壊したと思います。
現代のAIDSは根本的な治療方法がない恐ろしい感染症であり、いったんウイルスに感染すると発症を押さえる抗ウイルス薬を投与し続けるしかありません。しかしAIDSに対してさえ治療なしで元気な人がいます(日経サイエンス・2012年10月号『エイズを発症しない人々』)。免疫がどう作用するかは人によってものすごく違うわけです。
これは決して「抗体に対して強く反応するのがいい」のではありません。強く反応すると、自己に対しても強く反応する可能性が高く、アレルギーになったり自己免疫疾患を発病したりする。あくまで免疫の作用の「多様性」が大切なのです。そしてこの多様性こそ人間の集団が今まで生き延びてきた理由だと強く感じます。特定の病原菌に対して、全員が死ぬか全員がケロッしているかという集団は、早々に死滅してしまうでしょう。
免疫システムと社会
以上の、免疫システムの特徴としてまとめた
◆冗長性
◆多義性
◆ランダム性
◆偶然性
◆多様性
というポイントは「自己組織化」していくシステムの特徴であり、それは社会における複雑な組織体のありようとそっくりだと感じました。複雑な組織体とは、大企業、大都市、国家などです。それらは環境の変化に対しては自らを変え、再組織化をしていかないと生き延びられません。自己組織化するシステムは、同時に「危うさを内包するシステム」です。しかしそれ以外に組織体を永続させる原理はない。そう感じさせられました。
多田氏の2冊の本は2001年、ないしはそれ以前の本です。最近の10年で免疫学はさらに発展しました。その一つとして、人間と共生している細菌が免疫と関係していることが分かってきました。以下にそれを紹介します。
マイクロバイオーム:細菌叢
『エイズを発症しない人々』の記事が掲載されていた日経サイエンス・2012年10月号に『マイクロバイオーム』の特集記事が載っていました。マイクロバイオームとは、人間の皮膚や消化系に住み付いている細菌群の総体を言い、日本語では細菌叢(そう)です。これらの細菌の中には人間に役だっているものがあることは昔から知られていましたが、最近になって人間の免疫もこれらの共生細菌と関係していることが分かってきたのです。
東京大学・服部教授の文章に従って、その事情を紹介しますと以下のようです。まず服部教授は細菌の数の多さ語っています。
人体に住み付いている細菌は「常在菌」と言い、一時的に体内に進入して感染症を引き起こす「病原菌」とは区別されます。常在菌は病原性を示しません。
胎児の発育する子宮は無菌状態ですが、赤ちゃんが産道を通る時、最初の常在菌が皮膚に付着します。それ以降、生活環境の中にいる各種の細菌が体内に入ります。つまり常在菌は人間の誕生とともに体に生息をはじめ、一生をともにします。
そしてこの常在菌の中には「人間の免疫システムの一部として機能している」ものがあることが分かってきたのです。
J ・アッカーマンという米国のサイエンス・ライターが、バクテロイデス・フラジリス(フラジリス菌)について解説しています。その内容を要約すると以下のようです。この中の「制御性T細胞」は多田富雄氏の本に出てこないのですが、免疫反応を押さえる役割を持ったT細胞で、1995年に大阪大学の坂口志文教授が発見しました。近年研究が進んでいます。
あたりまえですが、人間の腸は異物に満ちています。そこは免疫システムの戦いにとっての「最前線」です。その最前線で、腸内細菌の出すサイトカイン相当の分子(PSA)が、結果として腸炎などの炎症をくい止め、免疫システムの過剰反応を抑制する役割を果たしているわけです。
記事によると、人間の常在菌の分布状況は人それぞれで違います。一卵性双生児であっても、生まれてからの生活によって常在菌は違う。免疫系は一人一人違っていて多様性があると前に書きましたが、人間と常在菌の共生を考えると人間だけを前提とするよりもっと多様性があるのです。
それと同時にフラジリス菌の例は、抗生物質の投与で(結果として)常在菌を殺してしまったり、必要以上に「清潔な環境」で生活をすることが「よくない結果」をもたらす可能性を示唆しています。我々人類は、何か間違ったことをやって来たのかもしれないのです。
人間は自己の細胞と細菌(常在菌)の共生系です。細菌の数は、自己の細胞の数の10倍以上です。そしてこの共生は単にギブ・アンド・テイクというレベルを越えています。フラジリス菌の例で言うと、フラジリス菌のPSA分子を作る遺伝子が、人間のゲノム(遺伝子の総体)を補っていると考えたほうがよい。
もちろんこれらの常在菌は免疫だけに役だっているのではありません。たとえば食物の消化です。
服部教授は、「ヒトは、ヒトゲノムとヒト常在菌叢ゲノムから成り立つ超有機体である」というノーベル賞学者(ジョシュア・レーダーバーグ。米国)の言葉を紹介していますが、まさにその通りです。前回の No.69「自己と非自己の科学(1)」で、多田富雄氏の本に従って
と書きました。免疫とは「自己とは何か」を生物的に規定しているものです。そしてヒトは自分の細胞の数の10倍もある細菌と共存している。つまり生物学的には
なのです。その「拡大された自己」の本格的な研究はこれからです。そこから得られた新たな知見は、人間の生き方に新たな示唆をもたらしてくれるだろうと思います。

|

|
|
|
多田富雄 『免疫の意味論』 |
多田富雄 『免疫・自己と非自己の科学』 |
免疫システムの特徴
多田氏の2冊の著作が描く免疫システムを概観すると、それは幾つかの際だった特徴をもっていることに気づきます。免疫システムの特徴をキーワードで表すと以下のようになると思います。
| 自己組織化 |
免疫は、刻々変わる「自己」と「非自己」に対応してシステムのありようを変え、再組織化していきます。この「自己組織化」は、変化する環境に対応して「困難な」目標を達成するべく運命づけられたシステムの必然なのでしょう。
| 冗長性 |
前回の No.69「自己と非自己の科学(1)」の中の「免疫のプロセス」で、多田氏の本からそのまま引用してB細胞とT細胞が絡む免疫の過程を紹介しました。このプロセスは、かなり複雑でまわりくどく、また冗長だと感じられます。もっとシンプルにできないのか。
免疫学の歴史上有名な「クローン選択説」があります。オーストラリアのウイルス学者・バーネットが1957年代に出した免疫のしくみを説明する学説で、現代免疫学の根幹をなす重要な学説です。それをB細胞を念頭において模試的に描いたのが次の図です。
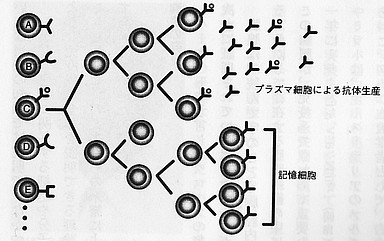
|
|
クローン選択説 (『免疫・自己と非自己の科学』より) |
図で左端のA・B・C・D・EはB細胞です。B細胞はそれぞれ抗原と特異的に反応する抗体を持っていて、それは細胞ごとに違う。ある抗原が図の「B細胞・C」の抗体と反応すると「B細胞・C」は分裂をはじめ、同じ遺伝的性質の子孫(=クローン)を増やしていく。それは「B細胞・C」のもつ抗体の大量生産につながり、抗原は駆除される。しかし一部のB細胞は組織内に長く残り、それが記憶細胞となって二次免疫反応を起こす・・・・・・。
バーネットが「クローン選択説」を出したときにはT細胞もB細胞も分かっていませんでした。しかし免疫の仕組みの基本的なところを見事に言い当てている。もうこの図で十分だという気がします。
しかし実際の免疫の仕組みはこのようにシンプルではありません。もっと複雑で冗長です。それは「免疫のプロセス」で引用したように、
| ① | B細胞による抗原の認識 | |
| ② | B細胞(APC)による抗原の取り込み | |
| ③ | ペプチドに分解した抗原をMHCに提示 | |
| ④ | T細胞による「抗原ペプチド・MHC複合体」の認識 | |
| ⑤ | T細胞の指令によるB細胞の増殖 | |
| ⑥ | T細胞の指令によるB細胞のプラズマ細胞への分化 | |
| ⑦ | プラズマ細胞による抗体の大量生成 |
というプロセスをたどります(キラーT細胞は省略)。「B細胞の抗体が抗原と反応しただけでB細胞が分裂を開始し抗体が大量生産される、ということはない」のです。なぜこんなにまわりくどくて冗長なのか。
おそらく上図のようなシンプルな仕組みは「あぶない」のだと思います。もし、自己のタンパク質と反応する抗体を持つB細胞があるとすると、その抗体が大量生産され、自己の破壊につながる。免疫の仕組みの冗長さは、それなりの理由があると考えられます。
| ランダム性 |
抗原を特異的に認識したり無力化するのは、T細胞抗原受容体(TCR)と免疫グロブリン(Ig)でした。そして、無限のバリエーションがある抗原を「特異的」に認識する仕組みは、まず遺伝子のランダムな組み替えで予期しえない TCR や Ig が作られ(利根川博士の発見)、そこから自己と反応するものが排除されるというプロセスでした。
免疫システムの根幹のところには、この「ランダム性」があります。非常に無駄が多い仕組みに見えますが、不測の事態(抗原の進入)の備えるにはこれしかないのでしょう。
| 偶然性 |
B細胞が作り出す抗体(免疫グロブリン。Ig)は、抗原を直接に認識します。しかし抗体の大量生産のトリガーを引くのはヘルパーT細胞であり、ヘルパーT細胞が認識するのは、MHCに提示された抗原の断片(9個のアミノ酸からなるペプチド)でした。
ここで誰しも思うのは「わずか9個のアミノ酸で抗原が確実に認識できるのだろうか」という疑問です。タンパク質はふつう数百のアミノ酸から成り、数千のアミノ酸で構成されるタンパク質も稀ではありません。9個で大丈夫なのか。
実際、多田氏の本には次の意味の記述があります(免疫の意味論)。
| ◆ |
自然界に存在するタンパク質を人工的に9個のアミノ酸からなるペプチドに分解しても、MHCの溝にうまく入り込むようなペプチドは著しく少ない。 | |
| ◆ |
立体構造も機能もお互いに全く無縁なタンパク質の間でも、しばしば共通のアミノ酸配列が見い出される。そういう部分が切り出されると、異なったタンパク質であるにもかかわらず同じ情報として認識されてしまう。 | |
| ◆ | どんなに人間にとって異物であっても、ペプチドまで分解されると人間の成分と区別できなくなる場合がある。マラリヤや住血吸虫に対して免疫が起こりにくい理由の一つがこれである。 |
抗原のタンパク質断片でもMHCの溝に「提示」可能なものは少なく、かつそれが人間のタンパク質と異なっているとは限らない・・・・・・。免疫システムは、ある種の「偶然性」の上に成り立っていることになります。多田氏が書いているように、この免疫システムの「欠陥」をついてくる「非自己」もある。
しかし多くの場合はこれで免疫が成り立っています。つまり、抗原もMHCもT細胞も「数が圧倒的に多い」ということなのでしょう。個々の確率は少なくても、母数が多いと安定的に作動するということがあります。
| 多義性 |
前回、No.69「自己と非自己の科学(1)」の「免疫のプロセス」に出てきたインターロイキン6(IL-6)というサイトカインがあります。これはヘルパーT細胞が作りだし、B細胞がプラズマ細胞に分化するように誘導する重要なサイトカインでした。しかしIL-6には別の作用もあるのです。多田氏の本によると、たとえば
◆キラーT細胞の分化を促す
◆マクロファージの働きを高める
などです。このあたりは免疫細胞に関係した作用であって、抗原を排除するのに役立つのだなと納得がいきます。ところが、IL-6は免疫とは直接関係のない作用もあるのです。つまり
◆神経細胞の成長を促す
◆骨を破壊する破骨細胞を作り出す
◆造血幹細胞や血小板を増やす
などです。同じ分子(タンパク質)が、こういった多種の作用を兼ねていいのかと思うのですが、それだけではありません。IL-6を作るのはヘルパーT細胞だけでなく、
◆マクロファージ
◆B細胞
◆皮膚の上皮細胞
◆血管の内皮細胞
でも作られる、とあるのですね。
B細胞をプラズマ細胞に分化させるという、免疫システムにとっては決定的に重要なサイトカインが、多数の別の「意味」に使われ、しかも各種の細胞がそのサイトカインを出す・・・・・・。IL-6というサイトカインはきわめて「多義性」があることになります。そしてこの多義性は IL-6 だけではないのです。
こんなことで免疫が成立するのかと心配になりますが、一応成立している。こういった「多義性」を含みつつ複雑な目的を実現するところに、自己組織化がポイントである免疫システムの特徴があるように感じます。
| 危うさを内包するシステム |
免疫システムは「危うさ」をもったシステムです。たとえば「胎児を母親の免疫システムから守る仕組み」です。前回の No.69「自己と非自己の科学(1)」で、多田氏の本に従って、
| 胎児は母親にとって異物だが、母親の免疫システムに攻撃されないのは胎児が母親と繋がっている胎盤の細胞表面からMHCが消える仕組みがあるからだ |
という意味のことを書きました。一見これで良いようなのですが、これは非常に「あぶない」仕掛けです。つまり人間には「MHCの発現を抑制する」遺伝子があり、胎盤だけでその遺伝子が働いているというようなことだと想像します。そうだとしたら、もし、がん細胞がその遺伝子を「利用」したらどうなるのか。免疫システムはがん細胞を攻撃できなくなります。事実、多田氏の本には「がん細胞は細胞のMHCを少なくしT細胞の監視からのがれる」という意味のことをが書かれているのです。
そもそもキラーT細胞は自己の細胞を殺す役割の免疫細胞です。もちろん殺すのは「細胞表面のクラス1・MHCに非自己(抗原)の断片を提示している自己の細胞」で、内部になんらかの異変が起きていると想定できる細胞です。ウイルスは自己に潜む非自己、ないしは自己に偽装した非自己なので、ウイルスの駆除までを目的とする免疫システムでは自己の細胞を殺す必要があるのは理解できます。
しかし免疫の仕組みには「偶然性」や「ランダム性」が内在しています。もし間違って本当の自己を提示している細胞を殺してしまったらどうなるのか。そういった反応性をもつキラーT細胞が「自己に反応する免疫細胞を抑制する仕組み(免疫的寛容)」を乗り越えて大増殖したらどうなるのか。免疫システムが自己を攻撃する事態が想像されます。
さらに想像すると、人間のDNAにある遺伝子は全部が活用されていわけでも何でもなく「眠っている」遺伝子がたくさんあることは生命科学の教えることです。その眠っている遺伝子が何かの拍子に発現し、通常では作られないタンパク質が細胞内で生産されるとどうなるのか。人間の免疫系はその細胞(自己)を攻撃することになります。
いろいろと心配の種が尽きない「危うさ」を内包しているのが免疫システムです。そして自己免疫病と総称される病気はまさにこの「危うさ」が現実のものとなった病気だと感じられます。
現代における難病のほとんどは「自己免疫病」です。たとえばバセドー病は甲状腺に反応する抗体ができる免疫反応で起こります。若年性糖尿病はインシュリンを生産する膵臓のβ細胞を自己の免疫系が破壊してしまうことで起こります。人間のほとんどの臓器・部位に対して、そこを攻撃してしまう自己免疫病があると言います。また体内の関節が炎症を起こし萎縮していくリウマチ性関節炎も自己免疫病です。付け加えますと、いわゆるアレルギーは抗原に対する免疫システムの過剰反応が原因です。それによって人間にとっては不要な、時には命にかかわりかねない炎症が起こる。
自己免疫病で最も破壊的なのはSLE(全身性エリテマトーデス)という難病で、これは自己のDNAに対する抗体ができてしまう病気なのです。DNAはすべての細胞にあるわけですから、免疫システムが「すべての自己」を攻撃する。攻撃されて破壊された細胞からはDNAが飛び出し(=抗原!!)、それがまた抗体の生産を促して、悪循環が際限なく続くことになります。
「自己免疫病」を典型とする免疫システムの「危うさ」は、非自己を排除するという極めて困難な目的の達成のために必然的に組み込まれてしまったものだと思いました。花粉症はその「ささやかな」例なのでしょう。
| 多様性 |
ここで言う「多様性」は、人間が個体ごとに違うという意味での多様性です。
免疫が発動するプロセスは、まず人それぞれに違うMHCが細胞表面にあり、そのMHCに抗原の断片が提示され、その「MHC・抗原断片の複合体」をT細胞が認識する、という過程でした。そこからロジカルに導かれる結論は、
| 免疫作用は人それぞれに違っている |
ということです。特定の抗原を認識しやすい人とそうでない人がいる。インフルエンザが流行して学校が閉鎖になっても、まったく病気にならない子もいる。逆にワクチンを接種してもそれが利かずにインフルエンザにすぐかかってしまう子もいる。スギ花粉を吸い込むとアレルギー症状を起こす人もいれば、起こさない人もいる。抗原(病原体)に対する反応は人によって非常に違うわけです。
世界史をみると15世紀のヨーロッパではペストが大流行しました。人口の3分の1が死亡したと言います。3分の1が死亡するという事態は大パニックを起こしたでしょう。しかしペストが大流行しているさなかでケロッとしている人も多いわけです。当時は「細菌による伝染病」という概念は全くありません。死んだ患者と接触した人も多かったはずです。それでも3分の2は死ななかった。もし人口の9割が死んだとしたらヨーロッパ文明は崩壊したと思います。
現代のAIDSは根本的な治療方法がない恐ろしい感染症であり、いったんウイルスに感染すると発症を押さえる抗ウイルス薬を投与し続けるしかありません。しかしAIDSに対してさえ治療なしで元気な人がいます(日経サイエンス・2012年10月号『エイズを発症しない人々』)。免疫がどう作用するかは人によってものすごく違うわけです。
これは決して「抗体に対して強く反応するのがいい」のではありません。強く反応すると、自己に対しても強く反応する可能性が高く、アレルギーになったり自己免疫疾患を発病したりする。あくまで免疫の作用の「多様性」が大切なのです。そしてこの多様性こそ人間の集団が今まで生き延びてきた理由だと強く感じます。特定の病原菌に対して、全員が死ぬか全員がケロッしているかという集団は、早々に死滅してしまうでしょう。
免疫システムと社会
以上の、免疫システムの特徴としてまとめた
◆冗長性
◆多義性
◆ランダム性
◆偶然性
◆多様性
というポイントは「自己組織化」していくシステムの特徴であり、それは社会における複雑な組織体のありようとそっくりだと感じました。複雑な組織体とは、大企業、大都市、国家などです。それらは環境の変化に対しては自らを変え、再組織化をしていかないと生き延びられません。自己組織化するシステムは、同時に「危うさを内包するシステム」です。しかしそれ以外に組織体を永続させる原理はない。そう感じさせられました。
多田氏の2冊の本は2001年、ないしはそれ以前の本です。最近の10年で免疫学はさらに発展しました。その一つとして、人間と共生している細菌が免疫と関係していることが分かってきました。以下にそれを紹介します。
マイクロバイオーム:細菌叢

| |||
東京大学・服部教授の文章に従って、その事情を紹介しますと以下のようです。まず服部教授は細菌の数の多さ語っています。
|
人体に住み付いている細菌は「常在菌」と言い、一時的に体内に進入して感染症を引き起こす「病原菌」とは区別されます。常在菌は病原性を示しません。
胎児の発育する子宮は無菌状態ですが、赤ちゃんが産道を通る時、最初の常在菌が皮膚に付着します。それ以降、生活環境の中にいる各種の細菌が体内に入ります。つまり常在菌は人間の誕生とともに体に生息をはじめ、一生をともにします。
常在菌のすみかは、口腔、鼻腔、胃、小腸・大腸、皮膚、膣など全身に及ぶ。人体にはおおよそ 1015 個(1000兆個)の常在菌が生息し、この数はヒトの細胞数(約60兆個)の10倍以上になる。常在菌の種類は1000種前後と見積もられている。 |
そしてこの常在菌の中には「人間の免疫システムの一部として機能している」ものがあることが分かってきたのです。
2010年には、自己免疫疾患を抑制する制御性T細胞の誘導に関係するバクテロイデス・フラジリスが、2011年には同様にこの制御性細胞を誘導するクロストリジウム属が発見された。 |
J ・アッカーマンという米国のサイエンス・ライターが、バクテロイデス・フラジリス(フラジリス菌)について解説しています。その内容を要約すると以下のようです。この中の「制御性T細胞」は多田富雄氏の本に出てこないのですが、免疫反応を押さえる役割を持ったT細胞で、1995年に大阪大学の坂口志文教授が発見しました。近年研究が進んでいます。
| ◆ | 腸内にすむフラジリス菌は表面にポリサッカライドA(PSA)という多糖類の分子をもっている。 | |
| ◆ | 樹状細胞はPSAを取り込み、未分化のT細胞に提示する。 | |
| ◆ | 未分化のT細胞はPSAに刺激されて制御性T細胞になる。制御性T細胞は炎症を起こす免疫細胞の活動を抑制して炎症を押さえる。 | |
| ◆ | PSAを持たないフラジリス菌は免疫細胞から攻撃されて生き延びることはできないが、PSAをもっていると免疫系からは攻撃されない。 |
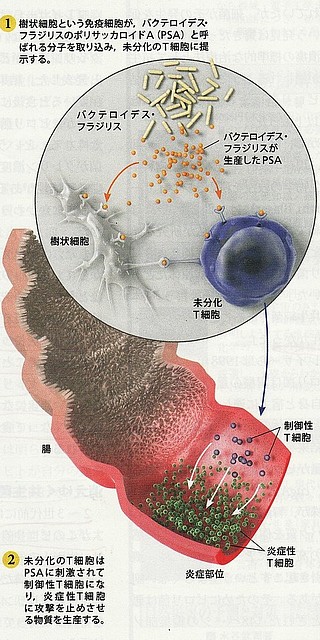
|
|
腸内細菌(フラジリス菌)が作り出す物質(PSA)がトリガーとなって 炎症反応が抑制される様子(「日経サイエンス」2012年10月号より引用) |
あたりまえですが、人間の腸は異物に満ちています。そこは免疫システムの戦いにとっての「最前線」です。その最前線で、腸内細菌の出すサイトカイン相当の分子(PSA)が、結果として腸炎などの炎症をくい止め、免疫システムの過剰反応を抑制する役割を果たしているわけです。
記事によると、人間の常在菌の分布状況は人それぞれで違います。一卵性双生児であっても、生まれてからの生活によって常在菌は違う。免疫系は一人一人違っていて多様性があると前に書きましたが、人間と常在菌の共生を考えると人間だけを前提とするよりもっと多様性があるのです。
それと同時にフラジリス菌の例は、抗生物質の投与で(結果として)常在菌を殺してしまったり、必要以上に「清潔な環境」で生活をすることが「よくない結果」をもたらす可能性を示唆しています。我々人類は、何か間違ったことをやって来たのかもしれないのです。
人間は自己の細胞と細菌(常在菌)の共生系です。細菌の数は、自己の細胞の数の10倍以上です。そしてこの共生は単にギブ・アンド・テイクというレベルを越えています。フラジリス菌の例で言うと、フラジリス菌のPSA分子を作る遺伝子が、人間のゲノム(遺伝子の総体)を補っていると考えたほうがよい。
|
もちろんこれらの常在菌は免疫だけに役だっているのではありません。たとえば食物の消化です。
有益な微生物の代表例は、バクテロイデス・テタイオタオミクロンだ。炭水化物を分解する能力が非常に優れていて、多くの植物性食品に含まれる大きな多糖類を、ブドウ糖などの小さくて単純で消化のしやすい糖類に分解できる。 |
服部教授は、「ヒトは、ヒトゲノムとヒト常在菌叢ゲノムから成り立つ超有機体である」というノーベル賞学者(ジョシュア・レーダーバーグ。米国)の言葉を紹介していますが、まさにその通りです。前回の No.69「自己と非自己の科学(1)」で、多田富雄氏の本に従って
| 免疫は自己と非自己(異物としての抗原)を認識し、非自己を排除することによって自己の統一性を保ち、生命を維持するしくみ |
と書きました。免疫とは「自己とは何か」を生物的に規定しているものです。そしてヒトは自分の細胞の数の10倍もある細菌と共存している。つまり生物学的には
| ヒトに常在している細菌は、自己の重要な一部 |
なのです。その「拡大された自己」の本格的な研究はこれからです。そこから得られた新たな知見は、人間の生き方に新たな示唆をもたらしてくれるだろうと思います。
2012-11-30 22:12
nice!(1)
トラックバック(0)



